Twin Prime Editing Mediated Deletion of CAG Repeats in the Mutant Huntington Gene
Thomas Batchelor
11-15-22
Twin Prime Editing Mediated Deletion of CAG Repeats in the Mutant Huntingtin Gene
Thomas Batchelor
Nov. 15, 2022
Background
Huntington’s Disease (HD) is an autosomal dominant progressive neurodegenerative disease. HD is characterized by deterioration of physical and cognitive abilities up to the point of death. The disease is caused by an expansion of the polyglutamine in exon 1 of the Huntingtin gene (HTT) on chromosome 4. Wild-type huntingtin protein is essential for a wide range of intracellular functions. The mutant protein disrupts normal cellular function. This leads to regression of neuron cells which leads to neurodegeneration. Individuals with >40 CAG repeats will develop debilitating symptoms over the course of their life. Currently, there are no FDA approved treatments for Huntington’s Disease. There are gene therapies in the works such as Uniqure’s that utilize a miRNA delivered via an AAV to silence the mutant HTT. However, few genome editing therapies are being developed for HD and there are currently none that utilize prime editing.
Prime editing (PE) is a precise genome editing method utilizing a derivative of the CRISPR-Cas9 system. PE was developed by Andrew Anzalone and David Liu at the Broad Institute. The first paper describing the mechanism was published in 2019. This technique uses a reverse transcriptase fused to a Cas9 nickase protein that, with the help of a modified guide RNA (pegRNA), targets a specific location in the genome for editing. Once the ssDNA cut is initiated, the reverse transcriptase uses the RNA template on the pegRNA to elongate the cut strand with the desired edit. This mechanism is displayed in figure 1.

Fig. 1: Prime Editing induced ssDNA cut and Edit Integration. https://www.nature.com/articles/s41596-022-00724-4
Aside from the ability to integrate a template-induced edit into the genome, PE has the added advantage of requiring three DNA hybridization steps before an edit can be integrated. This has the benefit of reducing off-target effects. In order to integrate an edit, the following three steps have to occur.
- Target DNA - pegRNA Spacer Complementarity
- Target DNA - pegRNA PBS Complementarity
- Target DNA - RT Product Complementarity
Following integration of the edit into the ssDNA as seen above there is “flap equilibration” between the 3’ edited and 5’ unedited DNA flap. Cellular degradation of the 5’ flap and ligation of the edited 3’ flap into the genome by DNA repair or replications results in stable installation of the edit. Mismatch Repair (MMR) is the only DNA repair mechanism that will always revert the edit back into the wild-type sequence.

Fig. 2: Flap Equilibration and Edit Integration Mechanism
https://www.nature.com/articles/s41596-022-00724-4
This research has been greatly expanded upon since 2019 to include twinPE. This uses the same mechanism as described above but with two separate pegRNAs to target both sides of a gene of interest. TwinPE can precisely insert or delete hundreds of base pairs of DNA. Figure 3 shows that the pegRNAs target opposite strands. Once cut, the reverse transcriptase utilizes the template to extend the 3’ flaps of both strands. These templates are complementary to each other so that the edited strands will bind. Excision of 5’ flaps will lead to integration of the desired edit.

Figure 3: Twin Prime Editing System for Replacement of DNA Between Cut Sites
https://www.nature.com/articles/s41587-021-01133-w
Great improvements have been made to optimize the editing efficiencies of Prime Editing. Changes have been made to the pegRNA to stabilize and protect the 3’ ends as well as improve the editor expression and nuclear localization. Twin Prime editing for the replacement of a 90bp site with a 38bp edit had an editing efficiency of just over 80%. This was before the full optimization of the pegRNA as well. Editing efficiencies in post-mitotic mouse cortical neurons were completed in 2019 with the original prime editing system and achieved editing efficiency of 7.1% with 0.58% indels. Since then studies have not been completed in post-mitotic cells.
While researching and considering methods for treating Huntington’s disease, Prime Editing appeared to have the greatest potential. Prime editing has an advantage over other techniques because it does not require the use of HDR for integration of the edit. There have been mentions of using this method for the treatment of a variety of genetic diseases however from assessment of the current literature it seems that Prime Editing has not been specifically designed and applied to Huntington’s disease. The following will summarize the novel approach of twin prime editing for removal of the mutant gene and integration of the edited gene as a therapy for HD.
Application
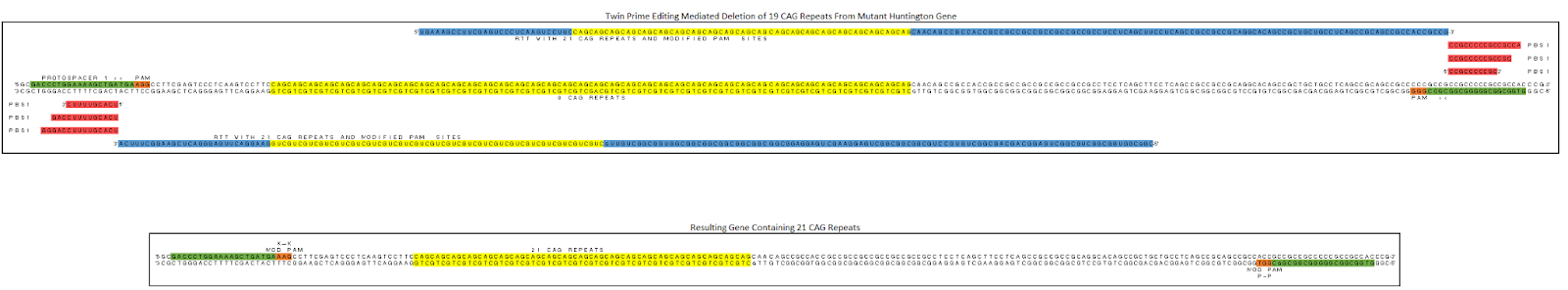
Fig 4. Design example of CAG removal using twinPE
The design follows the sample principle of the twinPE mechanism described above. Two pegRNAs have been designed to target opposite strands. The primer binding sites (red) are complementary to the strand that is being cut. Once each strand is cut the reverse transcriptase will use the template (blue) to create two complementary DNA strands that contain 21 CAG repeats. These complementary strands will hybridize and the natural DNA repair mechanisms will degrade the original strand and integrate the edited strand into the genome. The gene now encodes for a functional Huntingtin protein. The PAM sites are altered to silent mutations to ensure that the prime editor can not rebind to the gene.
- Protospacers (Green)
- PAM Sites (Orange)
- Reverse Transcription Primers (Red)
- Reverse Transcription Template (Blue)
There are numerous concerns when considering the optimization of this technique for in-vitro studies.
- Templates need to be made with thermodynamic and secondary structure considerations. The coding strand template in figure 4 has a GC concentration upwards of 80%. RNA secondary structures will also form which could prevent the reverse transcription. Other protospacers will need to be considered.
- Since the template only has 21 CAG repeats this means that it will always be shorter than the original gene. It is possible that there could be length limitations that would not allow the two edited strands to bind if the cell has excessive CAG repeats. This will have to be assessed by using cells with varying numbers of CAG repeats.
- The current design will cut both the mutant and non-mutant HTT on each chromosome. Since the PAM sites are altered it would only cut once. Ideally only the mutant is targeted. HD associated SNPs have been discovered which could present a solution to this issue.
It was proven in the first Prime Editing paper that primary cells can support prime editing. While editing efficiencies were significantly lower than in HEK293 cells, changes have been made to optimize the editing since then. This research would have two major benefits. It would specifically apply this new gene editing technology to Huntington’s disease for the first time and also assess editing in post-mitotic cells with the new optimizations.
Bibliography
https://www.nature.com/articles/s41586-019-1711-4
https://www.nature.com/articles/s41587-021-01133-w
https://www.nature.com/articles/s41596-022-00724-4